实验与药物语境下的 小干扰 RNA(small interfering RNA,siRNA)在序列上几乎总是以短双链形式出现:两条互补单链退火后,形成一条即将进入 RNA 干扰(RNA interference,RNAi)机器的 双链 RNA(double-stranded RNA,dsRNA)。若只记住「大约二十几个碱基」,仍难以解释为何同一靶序列的不同候选 siRNA 效率悬殊、为何 3’ 端多两个碱基、为何设计软件总强调 5’ 端热力学不对称。本篇把合成 siRNA 拆成可命名的结构区域,给出各区域常见的长度量级,并沿生命周期时间轴说明各区域在何时、通过何种分子接触发挥作用。
段末注释:siRNA 为长度约 19–25 bp(碱基对)量级的短 dsRNA,经 RNA 诱导沉默复合体(RNA-induced silencing complex,RISC)介导靶 信使 RNA(messenger RNA,mRNA)降解。
读前说明:下文默认讨论化学合成、经退火得到的经典不对称双链(最常见为每条单链 21 nt、中心 19 bp 配对、两端各 2 nt 3’ 突出端)。钝端双链、Dicer 底物(Dicer substrate,约 27 nt)及 shRNA(short hairpin RNA)茎区在文末对照;机制总览见 siRNA-01-生物学机制与功能原理.md,设计指标见 siRNA-05-设计指标脱靶与验证.md。
1. 整体分区:一条双链通常拆成几类结构?
从序列设计与蛋白结合两个视角,可把一条标准 siRNA 双链理解为 6 个功能区域(按引导链编号为主轴,过客链为互补镜像):
| 编号 | 区域名称 | 主要落在哪条链 | 常见长度量级 | 一句话功能 |
|---|---|---|---|---|
| A | 双链核心配对区 | 两链互补段 | 18–21 bp(最常见 19 bp) | 维持双链完整性、决定 Tm(解链温度)与递送中的稳定性 |
| B | 3’ 突出端 | 两链 3’ 端各一段单链 | 各 1–3 nt(最常见各 2 nt) | 模拟 Dicer 产物末端、参与 RISC 识别与抗外切酶策略 |
| C | 5’ 端区 | 引导链第 1 位及配对上下文 | 1 nt + 5’ 端热力学环境 | RISC 链选择、5’ 磷酸 与 AGO 结合口袋匹配 |
| D | 种子区 | 引导链第 2–8 位 | 7 nt | 靶搜索与 off-target(脱靶)敏感核心 |
| E | 中央区 | 引导链约第 9–12 位 | ~4 nt | 与 AGO2 介导切割在靶 mRNA 上的位点对应 |
| F | 3’ 端配对区 | 引导链约第 13–19 位(至突出端前) | ~7 nt | 提高 on-target 全配对亲和力,稳定引导链—靶复合物 |
区域 A、B 描述双链几何;区域 C–F 描述进入 RISC 后保留的引导链上的功能分段——二者在生命周期中时间重叠,但回答的问题不同(「双链是否完整」vs「哪段序列去识别靶 mRNA」)。
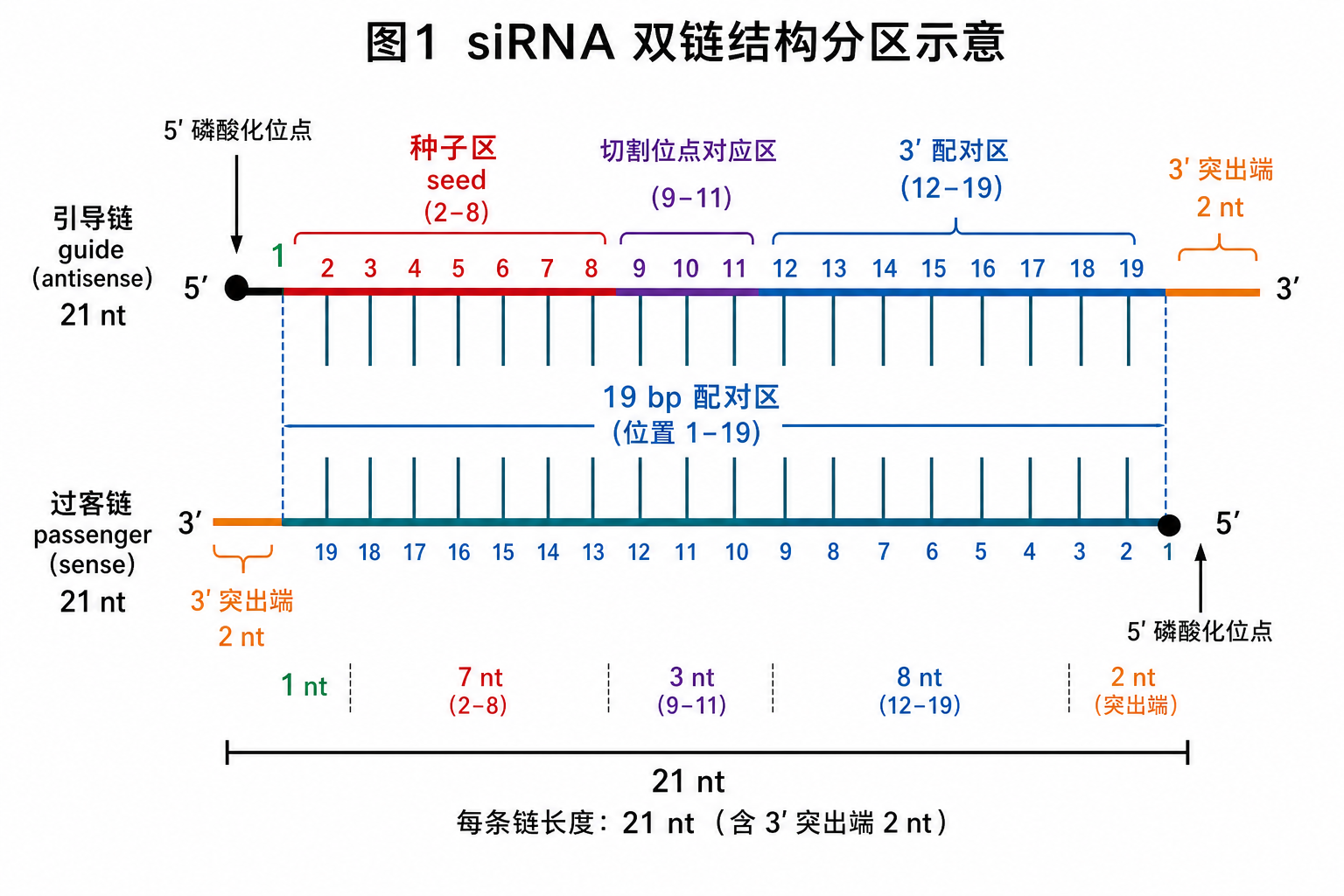
图 1 中,引导链(guide strand,反义链)通常对应靶 mRNA 的反义序列;过客链(passenger strand,正义链)在装载时被优先解离或切割。商业订单上的「sense / antisense」命名需与软件输出的 guide 对应关系核对,避免装反。
2. 双链核心配对区(A)
2.1 常见长度
- 经典设计:19 bp 连续 Watson–Crick 配对(A–U、G–C),两侧再各延伸 2 nt 单链突出,使每条单链总长 21 nt。
- 可接受窗口:配对区 18–21 bp 均有文献与产品实践;过短则双链不稳定、过长则接近触发先天免疫的 dsRNA 感知阈值,且未必提升敲低效率。
- 碱基对(bp)计数只计配对段;单链 nt(核苷酸)计数包含突出端。
2.2 生命周期中的功能
| 阶段 | 作用 |
|---|---|
| 合成与退火 | 决定双链 ΔG、Tm 与是否形成二聚体;GC 含量约 30–52% 为常见经验窗口(见 siRNA-05) |
| 血清 / 胞外 | 配对区越稳定,整体越抗部分核酸酶,但过稳可能妨碍 RISC 解链 |
| 内体 / 胞质 | 完整双链是进入装载复合体的底物形态 |
| RISC 装载 | AGO2 解旋双链时,配对区能量分布参与哪条链成为引导链(见 §4) |
| 靶切割后 | 引导链保留,配对区序列信息以单链形式继续参与靶识别 |
3. 3’ 突出端(B)
3.1 常见长度
- 标准:每条链 3’ 端 2 nt 非配对突出(两链突出序列可相同或互补,取决于设计)。
- 变体:1–3 nt 突出均有报道;0 nt(钝端)用于特定化学平台或 Dicer 底物经加工后产生的末端。
3.2 生命周期中的功能
- 生物真实性:天然 Dicer 产物常带 2 nt 3’ 单链突出;合成 siRNA 模仿该末端特征,有利于被 RISC 装载机器识别。
- 稳定性:突出端可进行 2’-O-甲基(2’-O-Me)等修饰而不直接阻断核心配对;过客链 3’ 端常用 脱氧(DNA)二聚体(如 dTdT)进一步降低核酸酶敏感性(属化学设计,非天然 RNA 序列)。
- 蛋白接触:AGO 家族 PAZ 结构域结合 3’ 端特征;突出端缺失或过长均可改变装载效率。
段末注释:PAZ 为 AGO 蛋白上结合 RNA 3’ 端的结构域模块。
4. 5’ 端区(C)与链选择
4.1 常见长度
- 严格说为引导链第 1 位核苷酸及其在双链 5’ 端的配对环境;讨论时常连同「哪条链 5’ 端更不稳定」一并考虑。
4.2 生命周期中的功能
- 化学状态:引导链 5’ 端常为单磷酸;未磷酸化可能影响人源 AGO2 装载。部分合成平台在 5’ 端预置磷酸或采用可转化化学基团。
- 热力学不对称规则(asymmetry rule):双链两端 5’ 稳定性不一致时,5’ 端配对更弱的一侧更易成为引导链——这是同一靶序列「正反链效率颠倒」的常见原因。
- RISC 装载后:5’ 端嵌入 AGO MID 结构域口袋,与第 2–8 位(种子区)共同构成「搜索前端」。
5. 种子区(D)
5.1 常见长度
- 引导链 5’ 端计数第 2–8 位,共 7 nt(第 1 位不计入经典 seed 定义)。
- 扩展讨论时偶见 2–9 或 2–10 的「扩展种子」用于脱靶预测,但算法默认值以 2–8 为主。
5.2 生命周期中的功能
| 阶段 | 作用 |
|---|---|
| 胞质靶搜索 | 与靶 mRNA 初步配对,决定结合速率与特异性 |
| 切割 | 对 siRNA 而言仍要求中央区与 3’ 区高度配对;种子单独不足以完成强效切割 |
| 脱靶 | 与非意图 mRNA 3’UTR(非翻译区)部分互补时,可产生种子驱动脱靶(详见 siRNA-05) |
miRNA(microRNA)与 siRNA 对比:miRNA 的调控更依赖种子不完全配对;siRNA 则强调全链互补,但种子仍是脱靶扫描的首要加权窗口。
6. 中央区(E)与切割位点
6.1 常见长度
- 引导链约第 9–12 位(不同综述对边界略有出入),对应双链约中心位置。
6.2 生命周期中的功能
- AGO2 的 slicer 活性在靶 mRNA 上切割的位置,与引导链第 10–11 位附近对齐(以 1 为 5’ 端起始计数的惯例)。
- 中央区错配会显著降低切割效率,即使种子配对良好。
- 设计时部分算法对中央区 AU 含量有偏好,但细胞类型与靶二级结构会掩盖统计规律。
若用位置变量表示,引导链长度 $L_g$ 时,种子区可写为 $i \in {2,\ldots,8}$,中央切割对应区常近似为 $i \approx 9$–$12$;此处 $i$ 为从 5’ 端开始的核苷酸序号。
7. 3’ 端配对区(F)
7.1 常见长度
- 引导链约第 13–19 位(19 bp 设计中为配对区 3’ 半段),直至 3’ 突出端之前。
7.2 生命周期中的功能
- 提供与靶 mRNA 的补充配对能量,稳定 RISC—靶复合物,尤其对 CDS(coding DNA sequence,编码区)靶向尤为重要。
- 与种子区共同决定 on-target 得分;全基因组扫描时,3’ 区错配惩罚常低于种子错配,但高于非种子区的随机错配。
- 过客链被释放后,该区以单链形式暴露,不再与互补链配对,直至结合靶 mRNA。
8. 生命周期总览:各区域何时「上场」
下列时间轴把 siRNA 从体外到降解的主要阶段与结构区域对应;细节机制见 siRNA-01、siRNA-00-前置知识-01-转染的生物学过程.md。
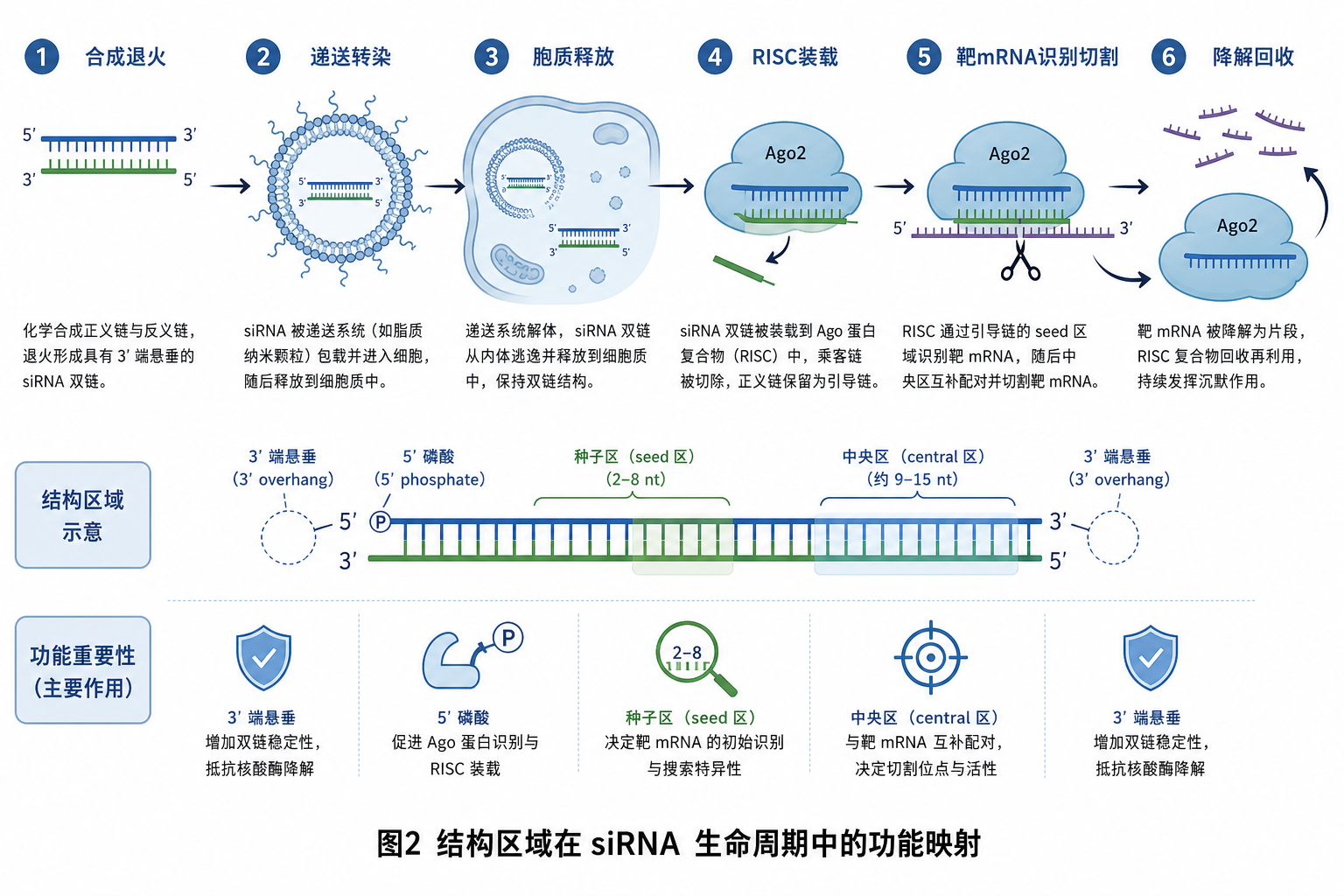
| 阶段 | 主导区域 | 要点 |
|---|---|---|
| 1. 合成、纯化与退火 | A、B | 配对区长度与 GC 决定双链是否形成;突出端序列影响后续修饰位点 |
| 2. 配制、血清与转染 | A、B(+ 化学修饰) | 完整双链抗降解;lipoplex / LNP(lipid nanoparticle)包裹与序列本身正交 |
| 3. 内吞与胞质释放 | 全链完整性 | 尚未按引导链分区;降解丢失任一端均降低有效浓度 |
| 4. RISC 装载 | C、B、A 热力学 | 5’ 不对称与 3’ 端识别决定 guide/passenger 命运 |
| 5. 靶识别与切割 | D、E、F | 种子启动搜索,中央区定位切割,3’ 区稳定全配对 |
| 6. 引导链周转 | D–F 单链形式 | siRNA 稀释、降解后效应衰减;蛋白半衰期独立于此 |
9. RISC 装载与切割:结构—蛋白接触示意
装载阶段是「双链语言」向「单链引导语言」的转换;切割阶段则几乎只谈引导链分区。
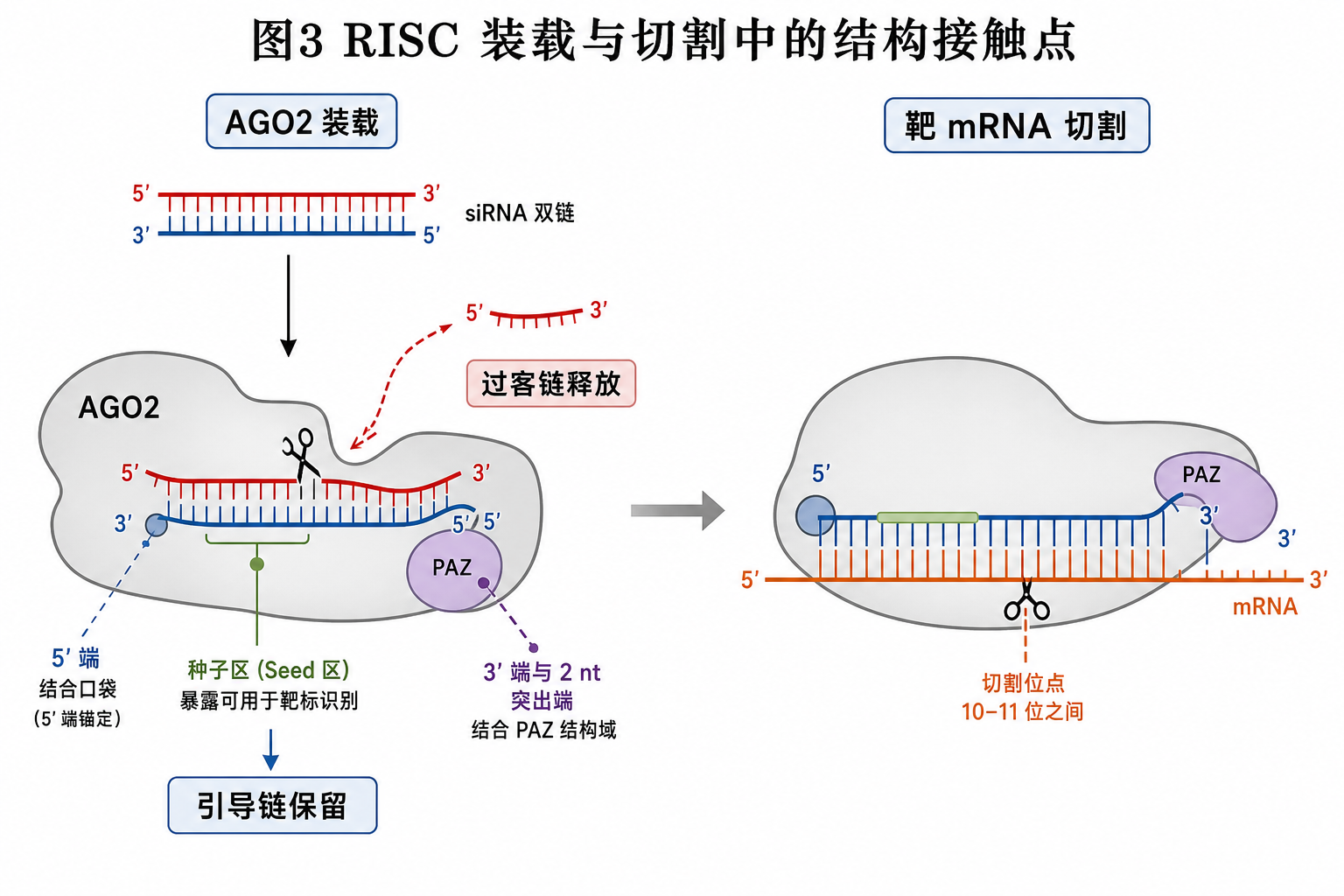
- 过客链:5’ 端热力学不利、构象不适合 AGO 通道时,被 AGO2 slicer 切割并释放。
- 引导链:5’ 端进入 MID 域,3’ 端(含突出端化学特征)接触 PAZ,种子与中央区朝向靶 mRNA。
- 靶 mRNA:与引导链形成双链段;切割发生在由中央区序列上下文指定的磷酸二酯键处。
10. 常见变体:长度与末端形态对照
并非所有有效沉默分子都符合「21 nt + 2 nt 突出」;理解变体有助于阅读 Dicer 底物与早期文献。

| 形式 | 配对区 / 单链长度 | 突出端 | 加工 | 典型用途 |
|---|---|---|---|---|
| 标准不对称双链 | 19 bp,链长 21 nt | 各 2 nt | 无需 Dicer 切割 | 实验室转染、多数药物化学 |
| 钝端 19-mer | 19 bp,链长 19 nt | 0 | 无 | 部分早期平台;装载效率常需优化 |
| 27-mer Dicer 底物 | ~25 bp,链长 ~27 nt | 各 2 nt | 胞内 Dicer 切出 21 nt 核心 | 提高装载不对称性的历史策略 |
| shRNA 茎区 | 设计为被 Dicer 识别 | 环与突出取决于载体 | 胞内转录 + Dicer | 稳定敲低、难转染细胞 |
shRNA 在细胞内被 Dicer 加工后,效应分子在结构上仍接近「带突出的短双链」,故上表 A–F 分区仍适用于成熟产物,只是其生命周期从 DNA 转录开始(见 siRNA-02-分子形式化学修饰与递送概览.md)。
11. 与序列设计表的交叉核对
下单或运行设计软件时,可用下列清单把「结构区域」落到具体碱基:
- 配对区是否 18–21 bp、GC 是否在合理窗口?
- 3’ 突出是否各 2 nt(或平台指定长度)?过客链是否需 dTdT?
- 5’ 端热力学是否有利于预期链成为 引导链?
- 种子(2–8)与 中央、3’ 配对区在全基因组脱靶扫描中是否高风险?
- 5’ 磷酸与 2’ 修饰位点是否按供应商规范标注?
小结
- 标准 siRNA 可按 双链几何(配对区 A、突出端 B)与 引导链功能段(5’ 端 C、种子 D、中央 E、3’ 配对 F)拆成 6 类区域。
- 最常见量级:19 bp 配对 + 各 2 nt 3’ 突出 → 单链 21 nt;种子 7 nt(第 2–8 位),中央约 4 nt,3’ 配对区约 7 nt。
- 生命周期上:A/B 主导合成稳定性与 RISC 入口识别;C 主导链选择;D–F 主导靶识别、切割与脱靶。
- 变体(钝端、27-mer、shRNA)改变的是加工路径,成熟效应分子仍 largely 共享上述分区逻辑。
相关文档
| 文档 | 关联内容 |
|---|---|
siRNA-01-生物学机制与功能原理.md |
Dicer、RISC、引导/过客链与切割 |
siRNA-05-设计指标脱靶与验证.md |
种子、GC/Tm、脱靶扫描 |
siRNA-02-分子形式化学修饰与递送概览.md |
3’ 修饰、递送与免疫 |
siRNA-00-前置知识-01-转染的生物学过程.md |
递送阶段(图 2 阶段 2–3) |