小干扰 RNA(small interfering RNA,siRNA)的 RNA 干扰(RNA interference,RNAi)活性与先天免疫(innate immunity)激活在分子上可以并存、分离或相互拮抗:同一条 siRNA 既可能高效装载 RISC(RNA-induced silencing complex)切割靶 信使 RNA(messenger RNA,mRNA),又可能在内体或胞质被模式识别受体(pattern recognition receptor,PRR)感知,诱导 I 型干扰素(type I interferon)与促炎细胞因子。后者会改变细胞转录组、翻译状态与存活率,使 qPCR/Western 读出难以区分「靶向沉默」与「免疫脱靶」,甚至出现敲低假象或敲低失败。
段末注释:PRR 为识别病原体相关分子模式的受体家族;I 型干扰素 主要包括 IFN-α、IFN-β,可广泛诱导干扰素刺激基因(interferon-stimulated gene,ISG)表达。
本篇聚焦序列依赖性免疫刺激(常称 immunostimulatory RNA,isRNA,与功能性 siRNA 相对);载体、浓度与 2’ 化学修饰的免疫学效应见 siRNA-02-分子形式化学修饰与递送概览.md。结构分区见 siRNA-06-序列结构区域与生命周期.md。
读前说明:免疫活性高度依赖细胞类型(树突状细胞 vs 普通贴壁细胞)、转染试剂、siRNA 浓度与是否 2’-O-甲基(2’-O-methyl,2’-O-Me)修饰;下文基序与阈值来自文献体外体系,宜作设计排除规则而非绝对定律。
1. 为何免疫反应会「干扰」敲低?
1.1 与 RNAi 平行的几条通路
| 传感器 | 主要配体特征 | 典型细胞区室 | 主要下游 |
|---|---|---|---|
| TLR7 | ssRNA、G/U 富集基序 | 内体(pH 酸性) | IFN-α(浆细胞样树突状细胞,pDC)、IFN-β |
| TLR8 | ssRNA、A/U 富集 | 内体 | IFN-α、TNF-α(单核细胞) |
| TLR3 | 长 dsRNA(通常 >~40 bp) | 内体 | IFN-β、IL-6 等 |
| PKR | dsRNA(长度依赖) | 胞质 | eIF2α 磷酸化 → 翻译全面抑制 |
| RIG-I / MDA5 | 5’ 三磷酸 ssRNA、长 dsRNA 等 | 胞质 | IFN-β、ISG |
标准 ~21 nt 双链在未修饰且经脂质转染进入内体时,TLR7/8 相关序列效应报道最多:双链在内体中部分解链后,U 富集单链上的短基序暴露,触发 MyD88—IRF7/NF-κB 轴(见 图 1)。
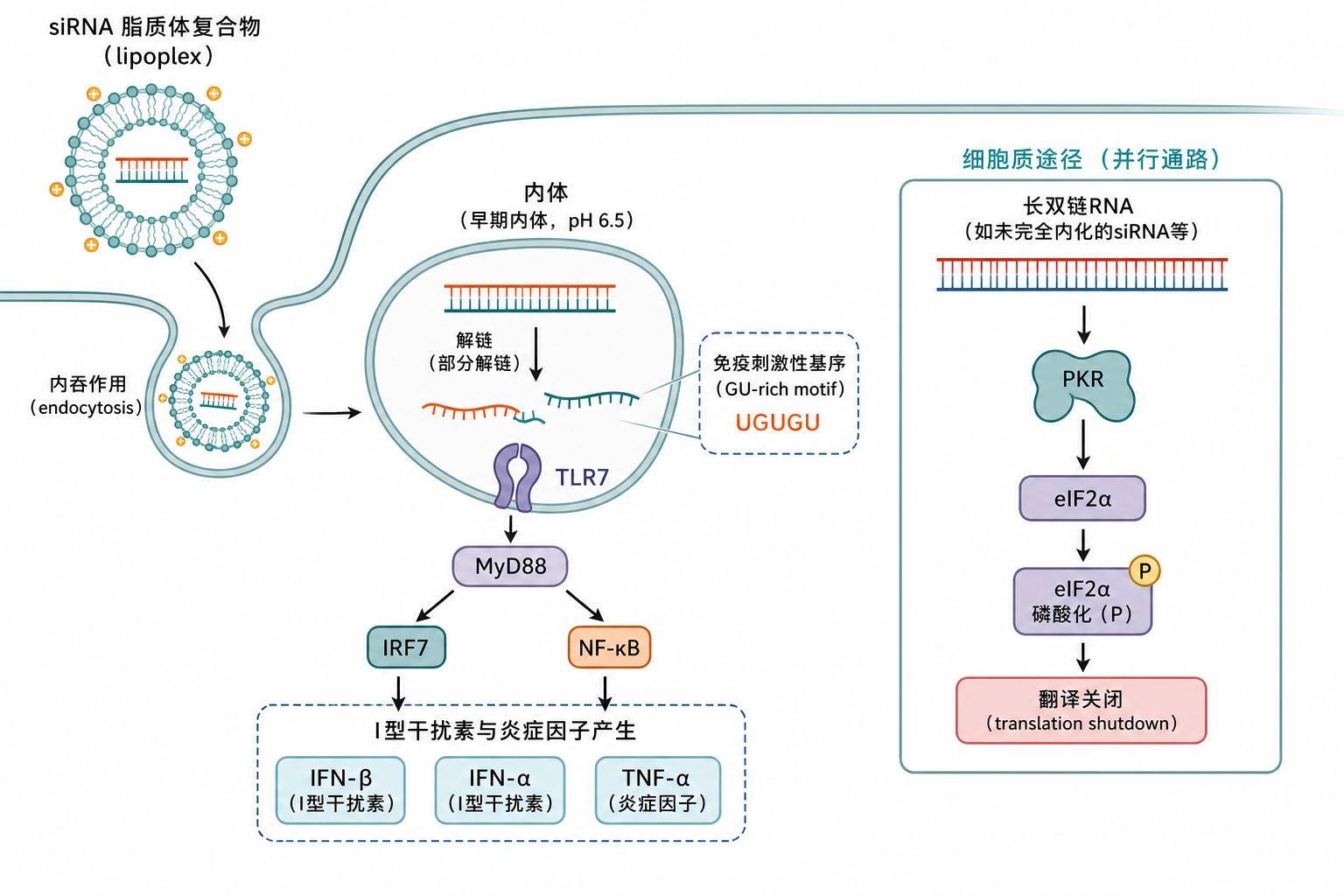
1.2 对敲低读出的具体干扰方式
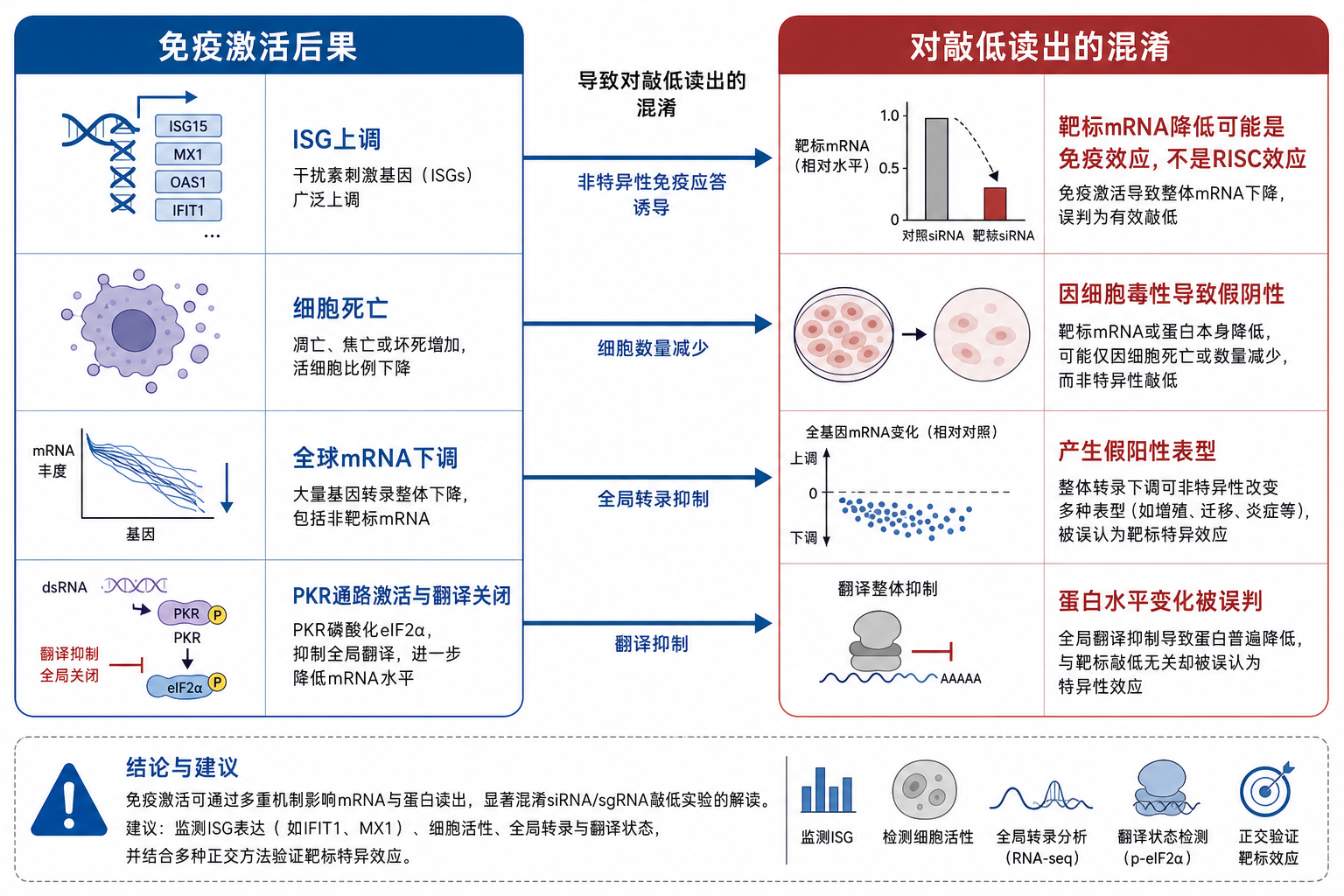
- 全局转录变化:ISG 上调;部分与靶基因无关的 mRNA 下降,qPCR 出现「非 RISC」的下降。
- 翻译关闭:PKR 或 IFN 通路抑制翻译,Western 条带普遍变弱,掩盖靶蛋白特异性变化。
- 细胞毒性:高浓度免疫刺激性 siRNA 导致死亡或生长停滞,有效 siRNA 浓度下降。
- 表型混淆:在抗病毒、炎症、凋亡通路研究中,表型可能来自免疫激活而非单基因敲低。
- 与脱靶叠加:免疫刺激性 siRNA 的转录组变化可与 seed(种子区)脱靶谱叠加,RNA-seq 解释难度上升(见
siRNA-05-设计指标脱靶与验证.md)。
段末注释:eIF2α 为翻译起始关键因子;其磷酸化可抑制整体蛋白合成。
2. 序列依赖性免疫刺激:主流机制模型
2.1 单链暴露模型(TLR7/8)
关键实验观察(人外周血单个核细胞,peripheral blood mononuclear cell,PBMC 体系):
- 同一 siRNA 的单链往往比退火双链更能刺激 TLR7/8。
- 双链 Tm(解链温度)越高、杂交越强,部分研究中免疫刺激反而减弱——提示内体中需有一定解链才能暴露刺激基序。
- U 含量更高的那条链更常被视为「免疫学意义上的敏感链」;在 U 富集背景中引入单个 G 可显著增强刺激(poly-U 上「点突变式」的 G)。
因此设计时不仅看「整条 G+U 百分比」(双链上 G+U 总数由长度固定),更要扫描单链上的局部基序与 U 连续段。
2.2 与递送、浓度的耦合
- 阳离子脂质 / lipoplex:促进内体摄取,TLR7/8 通路更易被观察到;裸转或电穿孔路径不同。
- 浓度:免疫刺激常呈剂量依赖;敲低亦依赖浓度——同一实验需设置阴性对照 siRNA(non-targeting control,NTC)与仅载体对照,且浓度矩阵一致。
- 化学修饰:2’-O-Me、2’-F 等可降低 TLR7/8 识别(见 §5);硫代磷酸(phosphorothioate,PS)主要改善稳定性,不自动消除序列免疫原性。
3. 常见免疫刺激性基序与文献锚点
下列基序多指单链 RNA 上下文(常为 siRNA 某一组分在内体解链后);位置多相对于该敏感链的 5’→3’。
3.1 基序速查表
| 基序 / 模式 | 示例(5’→3’) | 关联受体 / 效应 | 备注 |
|---|---|---|---|
| UGU 三联体 | …UGU… |
TLR7 | 最短报道之一;嵌入可提升刺激 |
| UGUGU 五联体 | …UGUGU… |
TLR7 | 经典强效基序;常用于 isRNA 设计 |
| GUCCUUCAA 九联体 | 完整 9 nt | TLR7、IFN-α | 见于强效 siRNA 正义链 3’ 端报道 |
| U 连续段 | UUUU(≥4) |
TLR7/8 | poly-U 背景敏感;偶发 G 增强 |
| GU 富集 / GU 二核苷酸重复 | GUGUGU…、GUGU 重复 |
TLR7 | 与 HIV TAR 等区域研究交叉 |
| AU 富集段 | AUAU…、高 A+U |
TLR8(单核细胞) | pDC 与单核细胞应答谱可不同 |
| UGGC 等四联体 | 文献列举扩展基序 | TLR7/8 | 综述中常与 UGUGU 并列 |
| 长 dsRNA 区段 | >30–40 bp 双链 | TLR3、PKR | 非典型 21 mer,但 shRNA、体外转录 需注意 |
并非「含 U 即禁用」:正常有效 siRNA 往往仍含多个 U;风险来自特定排列、单链暴露与实验系统敏感性的组合。
3.2 代表性参考文献
| 主题 | 文献 | 要点 |
|---|---|---|
| 序列依赖性 siRNA 免疫刺激 | Judge A.D. et al., Nat Biotechnol 2005;23:457–462. doi:10.1038/nbt1081 | 脂质配方 siRNA 可诱导 IFN;鉴定 UGU、UGUGU;可设计低免疫原性序列 |
| siRNA 激活免疫系统(双链感知) | Marques J.T. et al., Nat Biotechnol 2005;23:1399–1405. doi:10.1038/nbt1161 | siRNA 长度与结构对免疫的影响;与 Judge 同期互补 |
| TLR7 识别 ssRNA | Hornung V. et al., Nat Immunol 2005;6:933–940. | GU 富集 ssRNA 激活 TLR7 |
| TLR7/8 与 siRNA 序列决定簇 | Goodchild A. et al., BMC Immunol 2009;10:40. doi:10.1186/1471-2172-10-40 | 207 条 siRNA 筛选;UGUGU、GUCCUUCAA;U 计数与 Tm 相关性 |
| RNAi 与先天免疫综述 | Sioud M., Mol Ther 2005;11:605–616. | siRNA/isRNA「双刃剑」框架 |
| 免疫刺激性 RNA 综述 | Li L. et al., Front Immunol 2017;8:553.(PMC 综述) | 汇总 UGUGU、GUCCUUCAA 等 isRNA 基序与 TLR 通路 |
| 2’-O-Me 降低免疫原性 | Robbins M. et al., Mol Ther 2007;15:716–723. | 修饰 U 位点可减弱 TLR 识别、保留 RNAi |
| 修饰核苷酸与先天免疫 | Karikó K. et al., Immunity 2005;23:165–175. | 假尿苷 等修饰 RNA 降低 TLR 刺激(mRNA 疫苗前驱工作) |
| TLR7 依赖的 siRNA 刺激 | Heil F. et al., Eur J Immunol 2004;34:1826–1833. | U 富集 ssRNA 与 TLR7 |
设计软件 / 算法(用于 in silico 过滤,需以工具文档为准):
- DESiRNA、siDirect 等部分版本含免疫原性启发式评分;
- 商业合成平台常内置 GU/UGUGU 黑名单或 2’-O-Me 自动位点建议。
4. 实验上如何与「真敲低」区分
4.1 对照设置(最低配置)
| 对照 | 目的 |
|---|---|
| 仅转染试剂(mock) | 扣除载体本身炎症 |
| NTC siRNA(无关序列,同修饰) | 扣除转染与 siRNA 骨架效应 |
| 目标 siRNA | 待评序列 |
| (推荐)已验证低免疫 siRNA | 阳性沉默、低 ISG 的参照 |
转染条件(剂量、RNA/脂质 比、细胞密度)须在各组间完全一致。
4.2 常见评估方案
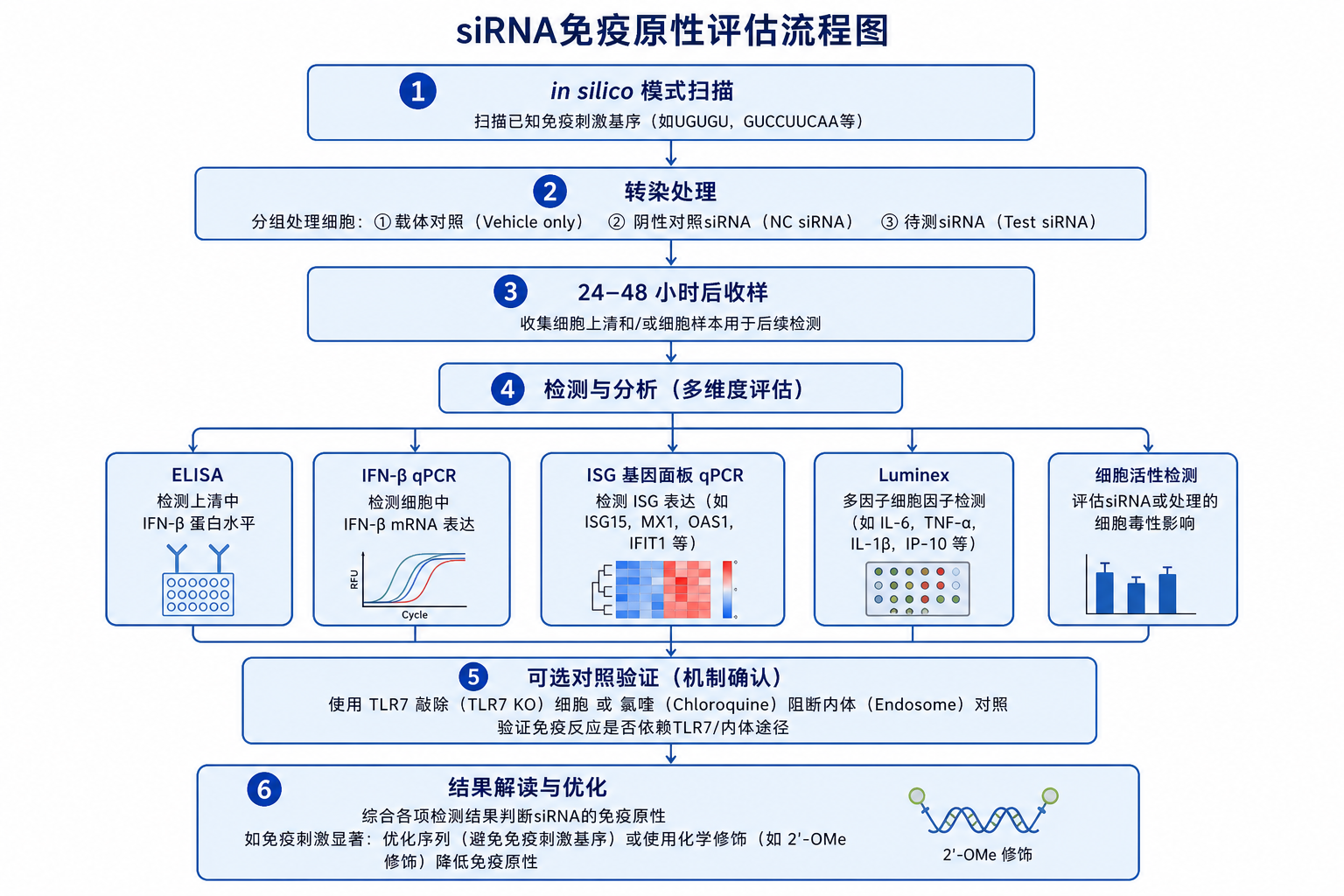
A. 分泌型读出(蛋白水平)
| 方法 | 靶标 | 优点 | 注意 |
|---|---|---|---|
| ELISA | IFN-β、IFN-α、TNF-α、IL-6 | 直观、定量 | 血清或上清需防细胞裂解污染 |
| Luminex / CBA 多因子 | 上述 + IP-10 等 | 一次测多种因子 | 成本较高 |
| 生物测定 | 上清处理 Reporter 细胞 | 敏感整合 IFN 活性 | 间接、需标化 |
B. 转录水平(ISG 面板)
常用 qPCR 基因(人/鼠需物种匹配):
- ISG15、IFIT1、IFIT3、MX1、OAS1、RSAD2(Viperin)、IRF7
时间点:转染后 6–24 h 常可见 ISG 上升;敲低读出若在 48–72 h,应同时检测早期免疫是否已发生。
C. 通路验证与机制对照
| 干预 | 预期若主要为 TLR7/8 |
|---|---|
| 氯喹 / 羟氯喹(内体酸化抑制) | 免疫读出下降(可能影响转染效率) |
| TLR7 敲低 / 敲除细胞或 PBMC 分型 | pDC 来源 IFN-α 减弱 |
| 单链 vs 退火双链平行 | 单链刺激 ≥ 双链 |
D. 组学层面
- RNA-seq:检查 ISG 模块是否在全转录组层面协同上调;与 off-target 基因列表交叉。
- 干扰素基因特征评分(interferon signature score):用于肿瘤免疫文献的 GSEA 基因集可借鉴。
E. 细胞活力与形态
- ATP 活力、LDH 释放、Annexin V/PI 流式;
- 免疫毒性导致的「敲低失败」常伴活力下降,而高效、低免疫 siRNA 可在较高浓度下仍维持活力。
4.3 结果解读简表
| 观测模式 | 更可能解释 |
|---|---|
| 仅目标 siRNA 致 IFN-β/ISG 升高,NTC 正常 | 序列依赖性 isRNA |
| 所有 siRNA 组均升高,mock 正常 | 转染过程或试剂批次问题 |
| mock 亦升高 | 血清、支原体、容器 RNA 污染或细胞系固有激活 |
| 靶 mRNA 下降且 ISG 暴升 | RNAi 与免疫并存;需 rescue 或换低免疫序列 |
| 靶 mRNA 不降但 ISG 高 | 免疫为主或 RISC 装载失败;检查序列与修饰 |
4.4 降低免疫原性的策略(与评估闭环)
- 重新选序列:避开 UGUGU、GUCCUUCAA 及长 UUUU;参考 Goodchild 2009 的 U 计数启发式。
- 2’-O-Me 修饰敏感 U 位点(尤其 passenger 链或 TLR 暴露面,依平台协议)。
- 降低剂量 在仍满足敲低的前提下做滴定。
- 换递送系统(可电离 LNP vs 传统阳离子脂质)——免疫谱不同。
- 药物开发中采用已批准的化学模式(GalNAc(N-乙酰半乳糖胺)偶联肝递送等)的平台经验,而非仅换序列。
5. 与系列其他文档的衔接
| 场景 | 建议阅读 |
|---|---|
| 设计阶段过滤免疫基序 | 本文 §3 + siRNA-05 §6 |
| 转染后仍无敲低 | 本文 §4 + siRNA-00-前置知识-01(内体逃逸) |
| RNA-seq 异常下调基因过多 | 本文 §4.2D + siRNA-05 §8–10 |
| 药物级安全 | siRNA-05 §12(非医疗建议) |
小结
- siRNA 可通过 TLR7/8(内体 ssRNA 基序)、TLR3/PKR(长 dsRNA)等通路激活先天免疫,与 RISC 介导的敲低相互独立又相互干扰。
- 常见高危基序包括 UGU、UGUGU、GUCCUUCAA、长 U 串及 GU 重复;权威序列筛选见 Judge 2005、Goodchild 2009。
- 评估应至少包含 NTC/mock 对照 + IFN-β 或 ISG 面板;机制验证可用内体酸化抑制或 TLR7 遗传学工具。
- 降低免疫原性需序列 + 修饰 + 递送 + 剂量联合优化,并以实验读出闭环,不能仅凭算法分数下单。
参考文献(精选)
- Judge A.D., Sood V., Shaw J.R., Fang D., McClintock K., MacLachlan I. Sequence-dependent stimulation of the mammalian innate immune response by synthetic siRNA. Nat Biotechnol. 2005;23(4):457–462.
- Marques J.T., Williams B.R.G. Activation of the mammalian immune system by siRNAs. Nat Biotechnol. 2005;23(11):1399–1405.
- Goodchild A., Nopper N., Tetzlaff M.T., et al. Sequence determinants of innate immune activation by short interfering RNAs. BMC Immunol. 2009;10:40.
- Hornung V., Guenthner-Biller M., Bourquin C., et al. Sequence-specific potent induction of IFN-α by short interfering RNA in plasmacytoid dendritic cells through TLR7. Nat Immunol. 2005;6(9):933–940.
- Sioud M. Induction of inflammatory cytokines and interferon responses by siRNA and isRNA: cellular mechanisms and potential clinical applications. J Mol Biol. 2009;386(5):1130–1138.(综述性讨论 siRNA vs isRNA)
- Robbins M., Judge A., Liang L., et al. 2’-O-methyl-modified RNAs act as TLR7 antagonists. Mol Ther. 2007;15(9):1663–1669.
- Karikó K., Buckstein M., Ni H., Weissman D. Suppression of RNA recognition by Toll-like receptors: the impact of nucleoside modification and the evolutionary origin of RNA. Immunity. 2005;23(2):165–175.